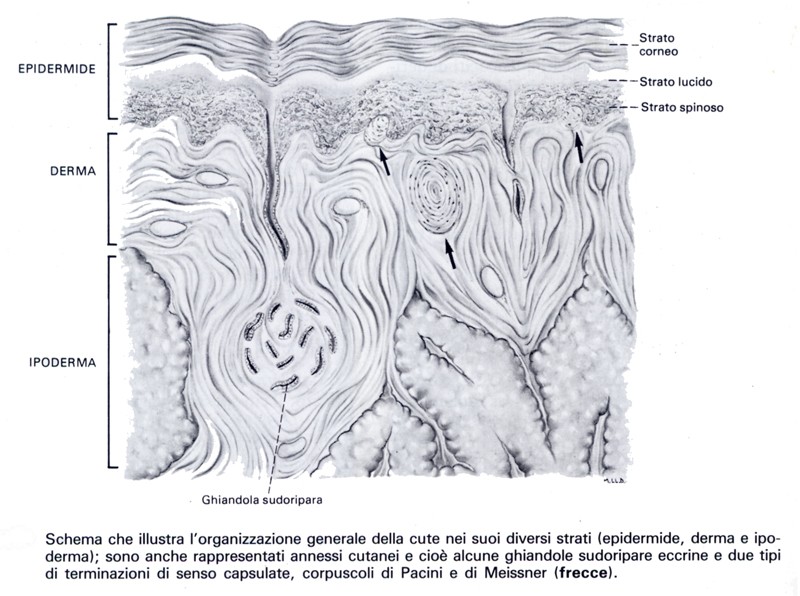
Adenoipofisi
L’adenoipofisi rappresenta il lobo anteriore dell’ipofisi. In essa si riconoscono una parte distale, una parte tuberale ed una parte intermedia; ognuna delle parti è caratterizzata da cellule che producono un determinato tipo ormonale.
Lo stroma che si trova fra le cellule ipofisarie e accompagna i capillari è di tipo reticolare e accoglie rari fibrociti e macrofagi. I capillari formano una rete molto ricca e sono di tipo sinusoidale, cioè di calibro non uniforme con dilatazioni e restringimenti che ben si adattano alla disposizione delle cellule parenchimali. Al microscopio elettronico la parete endoteliale presenta numerose fenestrature (o pori), chiusi da un diaframma. Una sottile lamina basale continua si trova all’esterno delle cellule endoteliali.
Indice
Parte distale
La parte distale dell’adenoipofisi è formata da cordoni di cellule, che vengono anche definiti come nidi cellulari, sostenuti da un delicato stroma reticolare e provvisti di una ricca rete di sinusoidi. Tralci fibrosi, derivati dalla capsula che avvolge l’organo, si insinuano nel parenchima ghiandolare, continuando con lo stroma più fine che compenetra e delimita i cordoni cellulari. Le cellule di questi cordoni hanno caratteristiche e dimensioni diverse.
Nei comuni preparati si distinguono, in base alle affinità tintoriali del citoplasma, tre tipi essenziali di cellule e cioè cellule cromofile acidofile, cellule cromofile basofile e cellule cromofobe. Le cellule cromofile appaiono tali per la colorabilità dei granuli presenti nel loro citoplasma.
Con le colorazioni tricromiche (tipo Azan-Mallory) è possibile distinguere, nell’ambito dei tre tipi fondamentali di cellule ipofisarie, alcuni sottotipi, quali le cellule α ed ε, facenti parte delle acidofile, le cellule β e δ, varietà delle basofile, e le cellule g, corrispondenti alle cromofobe. Un sesto tipo, le cellule h, sarebbe caratteristico dello stato gravidico.
Gli ormoni secreti dall’adenoipofisi, così come i granuli presenti nelle sue cellule, possono essere distinti in base alla loro natura di proteine semplici o di glicoproteine. Applicando il metodo di colorazione all’acido periodico-Schiff (PAS), che mette in evidenza i gruppi aldeidici presenti nelle glicoproteine, è stato possibile distinguere cellule ipofisarie PAS-positive, contenenti cioè granuli glicoproteici e cellule PAS-negative, i cui granuli sono invece costituiti da proteine semplici.
Cellule acidofile
Le cellule produttrici di ormoni proteici (ossia le cellule acidofile) sono ben riconoscibili per la loro colorabilità con i coloranti acidi e per i limiti cellulari netti che presentano. Sono PAS-negative e sono ritenute responsabili della produzione degli ormoni proteici. In condizioni normali rappresentano circa il 35% delle cellule epiteliali. È bene tuttavia sottolineare che i rapporti numerici tra i vari tipi di cellule sono variabili, sia in condizioni fisiologiche che patologiche.
- Le cellule α producono ormone della crescita (GH, o somatotropina, STH); hanno forma per lo più ovale, diametro massimo di 14-20 µm e presentano granulazioni piuttosto voluminose, di natura proteica, PAS-negative, colorabili con l’azocarminio. Al microscopio elettronico vi si osservano granuli di circa 300-400 nm di diametro, molto elettrondensi, sparsi in tutto il citoplasma, numerosi mitocondri con scarse creste laminari, un esteso apparato di Golgi e un ricco corredo di ribosomi sia liberi che aggregati a cisterne del reticolo endoplasmatico.
- Le cellule ε producono prolattina (PRL, o ormone lattogenico); hanno forma poliedrica e diametro pressoché uguale alle precedenti. Contengono granuli di notevoli dimensioni (600-900 nm), proteici, PAS-negativi, ben colorabili con l’orange G della miscela di Azan-Mallory. Al microscopio elettronico si riconoscono per il volume dei granuli e per il loro polimorfismo. Il citoplasma contiene inoltre uno sviluppato apparato di Golgi e un notevole reticolo endoplasmatico granulare. Queste cellule aumentano durante la gravidanza e l’allattamento.
Tra le cellule produttrici di ormoni proteici debbono infatti essere annoverate anche quelle che elaborano ACTH, cioè l’ormone adrenocorticotropo, che agisce sul trofismo della corteccia surrenale, in particolare della zona fascicolata, stimolandone l’attività secretoria che comporta una aumentata produzione di ormoni glicoattivi (cortisolo). Peraltro, se la natura proteica di questo ormone è un fatto ben accertato, altrettando non si può dire circa il tipo di cellule ipofisarie che lo producono. Secondo alcune osservazioni, trattasi di cellule acidofile, ma, in base ai dati emersi con il metodo degli anticorpi fluorescenti anti-ACTH, sembra trattarsi di cellule basofile (cellule B1). Le cellule, che con metodi immunoistochimici risultano contenere ACTH, reagiscono anche per un’altra molecola polipeptidica, chiamata β-lipotropina (β-LPH), che è un precursore del β-MSH e delle β-endorfine, polipeptidi questi ultimi implicati nel meccanismo di soppressione del dolore e largamente presenti in varie sedi del sistema nervoso centrale. Si ritiene pertanto che queste cellule, dette anche cellule ad ACTH-LPH, possano produrre tutti e tre i tipi di polipeptidi (ACTH, MSH, endorfine). Per le cellule ACTH-LPH è stata anche suggerita l’appartenenza al sistema APUD. Tuttavia, se alcune considerazioni d’ordine immunoistochimico e funzionale possono essere valide in questo senso, poco probabile o almeno non provata è la loro origine dalle creste neurali che è condivisa da tutte le cellule del sistema APUD. Il citoplasma delle cellule identificabili come corticotrope appare poco colorabile con i metodi della microscopia ottica e al microscopio elettronico risulta contenere granuli piuttosto scarsi e di modeste dimensioni (circa 200 nm). Bene sviluppati sono l’apparato di Golgi e il reticolo endoplasmatico granulare.
Cellule basofile
Le cellule produttrici di ormoni glicoproteici (ossia le cellule basofile) hanno per lo più forma rotondeggiante e volume maggiore rispetto alle altre cellule ipofisarie (circa 20-25 µm). Di contro, sono le meno numerose (circa il 15%) e hanno granuli piccoli, relativamente scarsi e quindi ben individuabili, che si colorano con il blu di anilina e con la reazione PAS. Esse posseggono un evidente apparato di Golgi e corrispondono alle cellule mucoidi di alcune classificazioni, in cui vengono così denominate per la natura glicoproteica del contenuto dei loro granuli.
A queste cellule viene attribuita la produzione degli ormoni glicoproteici ipofisari, ossia: ormone follicolo-stimolante (FSH), ormone luteinizzante (LH) e ormone tireotropo (o tireotropina, TSH).
Nell’ambito delle cellule basofile, PAS-positive, è stato possibile distinguerne alcune i cui granuli, oltre che PAS-positivi, sono positivi anche a colorazioni particolari come quella con paraldeide fucsina. Queste cellule, che si caratterizzano anche per particolarità di forma e di posizione, sono risultate responsabili della produzione dell’ormone tireotropo.
Le cellule gonadotrope sono PAS-positive, ma negative alla colorazione con paraldeide fucsina. È dubbio se, nell’ambito delle cellule gonadotrope, si possano distinguere quelle produttrici di FSH da quelle produttrici di LH. Dai dati recenti sembra tuttavia che si possano trarre le seguenti indicazioni:
Le cellule β producono l’ormone follicolo-stimolante; hanno forma prevalentemente ovale e sono localizzate soprattutto nelle porzioni laterali e posteriore della parte distale. Hanno granuli che si colorano intensamente in rosso con il PAS e con le tecniche miste (PAS più aldeide tionina o paraldeide fucsina). Al microscopio elettronico presentano granuli di circa 200 nm di diametro, dispersi in tutto il citoplasma, molto densi e aderenti alla membrana che li circonda. Le cisterne del reticolo endoplasmatico granulare sono caratteristiche in quanto appaiono dilatate e contenenti un materiale scarsamente opaco agli elettroni e di aspetto fioccoso.
Le cellule gproducono l’ormone luteinizzante; sono rotondeggianti, voluminose e hanno, rispetto alle precedenti, una posizione più centrale. I loro granuli assumono una colorazione rosso porpora con le tecniche combinate (PAS più aldeide tionina). Al microscopio elettronico presentano granuli di dimensioni costanti, intorno ai 250 nm, per lo più concentrati a un polo della cellula. L’apparato di Golgi appare meno sviluppato che nelle cellule follicolo-stimolanti e le cisterne del reticolo endoplasmatico granulare sono appiattite.
Le cellule δ producono l’ormone tireotropo; hanno forma irregolare, poliedrica, triangolare o spinosa e sono localizzate nella parte centrale della ghiandola. Sono positive al PAS, alla paraldeide fucsina, alla fucsina resorcina e all’aldeide tionina. Al microscopio elettronico presentano granuli di piccole dimensioni (circa 150 nm), disposti per lo più a ridosso della membrana cellulare. L’apparato di Golgi è generalmente concentrato a un polo del nucleo.
Dopo asportazione della tiroide si trasformano in elementi vacuolizzati, tipici, detti cellule da tiroidectomia.
Cellule cromofobe
Le cellule cromofobe sono le più numerose (circa il 40-50%). Hanno citoplasma relativamente scarso, poco colorabile, PAS-negativo e limiti mal definiti; nei comuni preparati si riconoscono come gruppi cellulari con nuclei ravvicinati e citoplasma poco distinto. Sono generalmente considerate come elementi di riserva, non funzionanti, da cui possono trarre origine, a seconda delle situazioni, gli elementi cromofili. Peraltro, molte cellule, interpretabili come cromofobe, sono in realtà elementi cromofili poveri di granuli. Questo fatto, mentre pone in discussione la consistenza numerica reale delle cellule cromofobe vere, fa sorgere anche il problema se queste cellule siano veramente elementi precursori delle cromofile e non piuttosto cellule cromofile degranulate o parzialmente degranulate a seguito di intensi fenomeni escretori. È possibile che ambedue le ipotesi siano attendibili, tenuto conto del fatto che le mitosi, nell’ipofisi adulta, sono molto rare e che quindi le diverse situazioni fisiologiche, implicanti la prevalenza funzionale momentanea di un tipo o di un altro di cellule in rapporto alla produzione di determinati ormoni, sono da porsi in relazione non tanto con variazioni dell’attività proliferativa, ma piuttosto con fluttuazioni nell’ambito della popolazione di cellule cromofobe.
Al microscopio elettronico si identificano come cellule cromofobe quelle povere di organuli (soprattutto apparato di Golgi piccolo e scarso ergastoplasma) e prive di granuli di secreto. Queste cellule agranulari hanno spesso forma molto irregolare (cellule stellate), perché fornite di prolungamenti citoplasmatici che si insinuano tra le cellule granulari, formando una specie di rete nella quale queste ultime sono comprese. Sottili prolungamenti si spingono inoltre negli spazi pericapillari, interponendosi tra la parete capillare e il citoplasma delle cellule granulari. Si ritiene che le cellule stellate possano avere una funzione di sostegno e possano anche intervenire attivamente nella composizione e circolazione dei fluidi interstiziali dell’ipofisi. Talvolta le cellule agranulari delimitano piccole cavità irregolari, simili a follicoli per cui sono state anche definite cellule follicolari. È stato anche suggerito che queste cellule siano progenitrici delle cellule adrenocorticotrope.
Più difficile, anche a livello ultrastrutturale, è l’identificazione di cellule in cui, a un certo sviluppo degli organuli citoplasmatici, particolarmente dell’ergastoplasma, si associa la presenza di pochi granuli di piccole dimensioni. Trattasi probabilmente di cellule in via di trasformazione in elementi cromofili. Se invece l’ergastoplasma è pressoché assente e il Golgi poco sviluppato, pur essendo presente qualche granulo di secreto, è più logico pensare che si tratti di cellule cromofile in avanzato stadio di degranulazione e in via di ritornare allo stato di elementi cromofobi.
Parte tuberale
La parte tuberale dell’adenoipofisi è la porzione del lobo anteriore che si applica sul peduncolo ipofisario, che contribuisce a formare. È costituita da cellule cubiche o cilindriche, disposte in cordoni inframezzati da una ricca rete vascolare facente parte del sistema portale ipofisario. Le cellule sono scarsamente colorabili, del tipo delle cromofobe e possono contenere nel citoplasma goccioline di sostanza colloide; talvolta si organizzano a formare piccoli follicoli e nidi di elementi piatti di tipo squamoso oppure palizzate di elementi che, peraltro, non manifestano una apparente attività secretiva.
Non si conosce la funzione della parte tuberale.
Parte intermedia
La parte intermedia dell’adenoipofisi aderisce alla neuroipofisi e può essere separata dalla parte distale dai residui della fessura ipofisaria. È costituita da cellule in prevalenza basofile, gruppi delle quali possono spingersi nella neuroipofisi. Molto spesso le cellule si dispongono a circondare cavità di tipo follicolare, contenenti una sostanza colloide più o meno intensamente colorabile.
La parte intermedia produce l’ormone melanotropo (o stimolante i melanociti, MSH, o intermedina). L’MSH è un polipeptide che presenta molte analogie nella sequenza degli aminoacidi con l’ACTH. Le cellule che producono MSH sarebbero sparse in tutto il lobo anteriore e sarebbero le stesse che producono ACTH.
Immagini
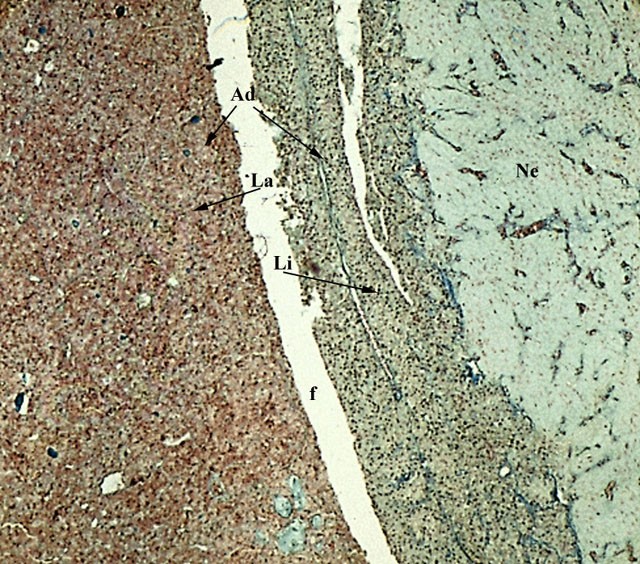
Ipofisi di uomo. Organo impari e mediano, appeso mediante il peduncolo ipofisario alla faccia inferiore del cervello: si trova pertanto nella scatola cranica entro la sella turcica dello sfenoide. E’ costituita da due parti distinte per struttura, funzione e origine embriologica: la neuroipofisi (Ne) situata posteriormente, fa parte del diencefalo, è costituita da fibre nervose amieliniche e da pituiciti, che sono cellule gliali. L’adenoipofisi (Ad) ha una struttura epiteliale e comprende il lobo anteriore (La), il lobo intermedio (Li) e la lingula. Negli individui giovani (fino al 16° anno) è possibile evidenziare la fessura ipofisaria (f) che divide il lobo anteriore da quello intermedio. (Colorazione Mallory Azan).
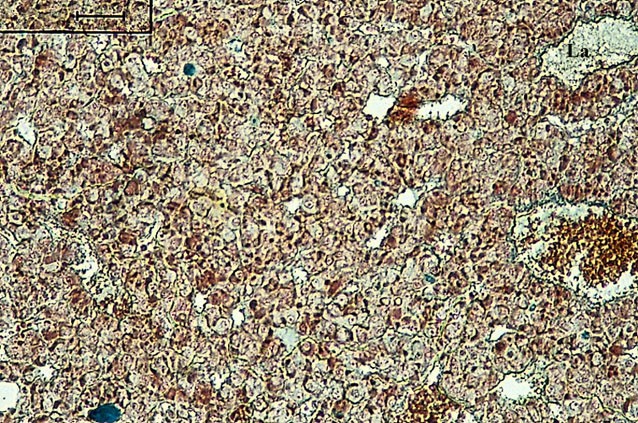
Particolare dell’immagine precedente visto a maggiore ingrandimento. (Colorazione Mallory Azan).
Il lobo anteriore (La) della adenoipofisi ha un colore più scuro ed è dato da cordoni di cellule epiteliali inframezzati da capillari sanguigni molto ampi. Tali cellule hanno numerosi granuli citoplasmatici che rappresentano il loro secreto e che le fanno classificare in acidofile (somatotrope, luteotrope, corticotrope), basofile (gonadotrope follicolostimolanti, gonadotrope luteinizzanti, tireotrope), cromofobe (cellule in stato di riposo).
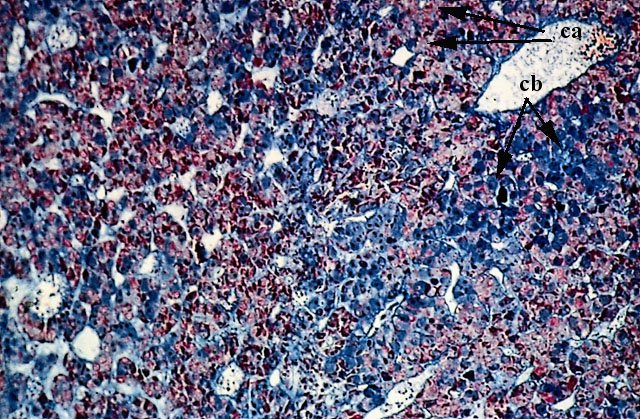
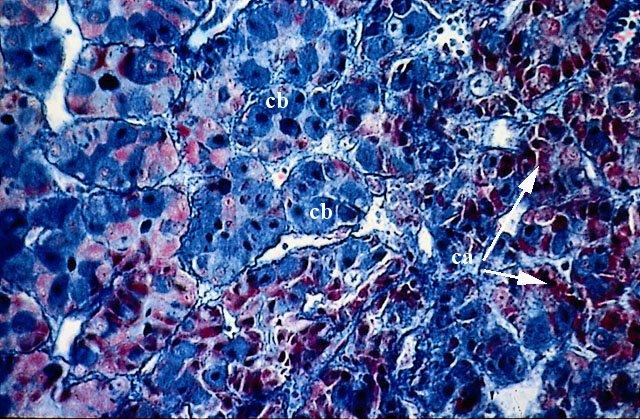
L’adenoipofisi appare costituita da cordoni epiteliali organizzati in nidi di cellule immersi in uno stroma reticolare. E’ riccamente vascolarizzata. (Colorazione Mallory Azan).
Gli elementi epiteliali che costituiscono i cordoni ed i nidi sono omogenei solo per la forma poliedrica, mentre appaiono molto variabili per dimensioni e caratteri strutturali.
Vi si distinguono diversi tipi cellulari che corrispondono ai diversi ormoni isolati dall’ipofisi stessa.
1. cellule acidofile (ca): secernono ormoni proteici (somatotrope, luteotrope, corticotrope).
2. cellule basofile (cb):secernono ormoni glicoproteici (gonadotrope follicolostimolanti e luteostimolanti, tireotrope).
3. cellule cromofobe sono probabilmente elementi indifferenziati che non presentano attività secretiva.