Epatociti
Gli epatociti hanno forma di poliedri con sei o più facce e misurano in sezione 20-25 µm. Sono disposti a formare lamine unicellulari tra loro anastomizzate e interposte al decorso dei sinusoidi.
Delle varie facce che gli epatociti presentano, alcune sono rivolte verso la parete sinusoidale (poli vascolari), altre sono relativamente pianeggianti e a stretto contatto con analoghe superfici di cellule epatiche adiacenti, insieme alle quali delimitano gli esili capillari biliari (poli biliari).
Le superfici delle cellule epatiche rivolte verso i sinusoidi sono provviste di piccole vescicole di pinocitosi e di corti ma numerosi microvilli. Questi si proiettano in uno spazio sottoendoteliale variabile per ampiezza e contenente filtrato plasmatico (spazio perisinusoidale di Disse). Gli spazi di Disse non risultano limitati soltanto all’area sottoendoteliale, ma si possono talvolta estendere anche tra gli epatociti adiacenti dove formano strette e irregolari fessure intercellulari (recessi perisinusoidali). In tutti questi spazi perisinusoidali come in quelli sottoendoteliali le superfici degli epatociti sono provviste di numerosi microvilli. Si realizza così un esteso sistema microlabirintico di spazi pericapillari e intercellulari dove si raccoglie e circola liberamente il filtrato plasmatico e dove vengono quindi favoriti gli scambi tra cellule epatiche e sangue. Il prodotto del filtrato sanguigno e delle attività metaboliche cellulari si presenta sotto forma di un liquido che, scorrendo all’interno degli spazi di Disse, viene convogliato verso la periferia del lobulo dove si versa nelle aree portobiliari, in uno spazio circoscritto da numerose fibre collagene e reticolari (spazio di Mall).
I liquidi contenuti negli spazi di Disse e di Mall non sono linfa in senso stretto. Sono fluidi interstiziali che contribuiscono alla formazione della linfa solo quando vengono drenati nei vasi linfatici terminali spesso presenti nelle stesse aree portobiliari, accanto agli spazi di Mall.
Le superfici cellulari che servono a delimitare i capillari di secrezione biliari (poli biliari dell’epatocita) sono pianeggianti e strettamente accollate, fatta eccezione per una piccola area in cui la superficie appare incavata a doccia. Questa, giustapponendosi a un’analoga depressione presente sulla parete della cellula adiacente, delimita la parete del capillare biliare. Le superfici dell’epatocito che delimitano il capillare biliare sono fornite di numerosi e brevi microvilli sporgenti nel lume e sono separate dagli spazi intercellulari adiacenti ad opera di complessi di giunzione (zone occludenti).
In condizioni normali nessuna comunicazione è possibile tra il polo biliare chiuso dai complessi di giunzione e il polo vascolare della cellula epatica. Pertanto, superfici adiacenti possono contemporaneamente essere impegnate nell’ambito dello stesso epatocito a svolgere funzioni di assorbimento e di secrezione di materiali completamente diversi. Soltanto in condizioni patologiche (ittero da stasi) gli spazi di Disse e i capillari biliari appaiono notevolmente dilatati e i complessi di giunzione risultano allentati per cui facilmente la bile può essere riversata negli spazi sottoendoteliali e da qui raggiungere il lume dei sinusoidi.
Nell’epatocito si trovano in abbondanza tutti gli organelli citoplasmatici oltre a un certo numero di inclusi caratteristici.
Gli organelli cellulari hanno una collocazione citoplasmatica piuttosto precisa, in genere legata a una specifica funzione dell’epatocito.
I lisosomi e i perossisomi (organuli di forma sferica ricchi in perossidasi e caratterizzati dalla presenza di un incluso cristallino) e soprattutto i sacculi e le vescicole del complesso di Golgi sono prevalentemente localizzati in corrispondenza del polo biliare dove sono impegnati in funzioni secretive. Il reticolo endoplasmatico si presenta nelle sue due forme tipiche, liscia e rugosa, mentre i ribosomi liberi (poliribosomi) sono anch’essi abbondantemente distribuiti nel citoplasma. Il numero e l’estensione dei canalicoli e delle cisterne del reticolo endoplasmatico dell’epatocito sono soggetti a continue variazioni che riflettono il diverso impegno funzionale dell’organo.
Per esempio, un rapporto topografico preciso è stato osservato tra inclusi di glicogeno e membrane del reticolo endoplasmatico liscio; tali membrane con il loro corredo enzimatico (glucoso-6-fosfatasi) possono contribuire al processo di glicogenolisi cui segue l’immissione di glucosio nel sangue. Inoltre, i lipidi assorbiti dal sangue attraverso il polo vascolare dell’epatocito verrebbero convogliati nel reticolo endoplasmatico liscio alle cui membrane si trova legata parte degli enzimi responsabili della sintesi del colesterolo e della degradazione di molti farmaci liposolubili (come i barbiturici). Il reticolo endoplasmatico granulare e i ribosomi liberi sono responsabili della sintesi delle proteine plasmatiche prodotte dal fegato e riversate in circolo attraverso il polo vascolare degli epatociti (albumina, fibrinogeno).
Al reticolo endoplasmatico degli epatociti spetta inoltre l’importante funzione dell’assemblaggio di molecole lipoproteiche che rappresentano una forma di smaltimento degli accumuli lipidici. Al loro polo vascolare gli epatociti liberano infatti, nello spazio di Disse, particelle denominate VLDL (Very Low Density Lipoproteins) che appaiono già all’interno dei tubuli del reticolo endoplasmatico, vicino al polo vascolare, come inclusi di bassa densità elettronica. Le VLDL sono complessi lipoproteici costituiti da un nucleo di trigliceridi, da un involucro costituito da un’apoproteina e da una miscela di colesterolo e fosfolipidi.
Al reticolo endoplasmatico liscio degli epatociti spettano inoltre altre importanti funzioni, come quelle detossificanti nei riguardi per esempio di farmaci (come i barbiturici). Queste funzioni si svolgono prevalentemente attraverso i processi di glucuronoconiugazione catalizzati da enzimi presenti nelle membrane del reticolo endoplasmatico.
I mitocondri, in genere molto numerosi, hanno forma tondeggiante e sono provvisti di lunghe creste; essi possono subire modificazioni di numero e di forma in rapporto alle speciali richieste funzionali della cellula epatica.
Infine, legati al metabolismo cellulare (o alla patologia dell’epatocito), possono trovarsi nel citoplasma inclusi di natura lipidica, oltre a pigmenti biliari, ferritina, lipofuscina.
Il nucleo è solitamente molto voluminoso e tondeggiante e contiene uno o più grossi nucleoli. In condizioni normali le cellule epatiche dell’adulto raramente entrano in mitosi, ma frequenti sono i casi di epatociti binucleati o forniti di nuclei con corredo poliploide. Soltanto nei processi rigenerativi le mitosi possono farsi numerose.
Sebbene tutte le cellule epatiche possano svolgere indistintamente le complesse funzioni dell’organo, il loro grado di attività sembra dipendere in certa misura dalla posizione che i singoli elementi occupano nel contesto del lobulo epatico.
Così, in un lobulo classico, le cellule epatiche in posizione più periferica si troverebbero in uno stato di elevata attività, quelle della zona intermedia avrebbero un’attività variabile e infine quelle disposte intorno alla vena centrolobulare sarebbero in uno stato di minima attività.
Questa diversa attività degli epatociti contenuti nelle tre zone funzionali in cui è suddiviso il lobulo dipende ovviamente dalla capacità che le cellule hanno di assumere ossigeno e sostanze nutritizie dal sangue circolante nei capillari adiacenti e proveniente, come già detto, dai rami terminali dell’arteria epatica e della vena porta localizzati alla periferia del lobulo. Pertanto le aree più periferiche del lobulo epatico risultano topograficamente più favorite, rispetto a quelle intermedie e queste ultime rispetto a quelle più centrali, a ricevere un maggior quantitativo di sangue arterioso.

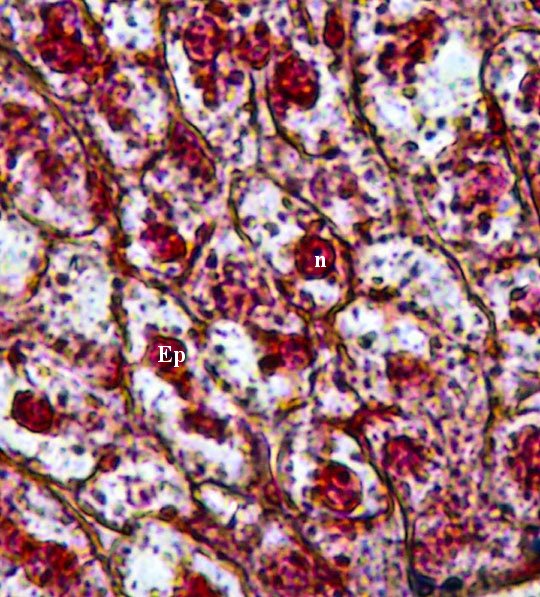
Particolare ingrandito dell’immagine precedente. (Colorazione Mallory Azan). Sono ben visibili le cellule epatiche (Ep), con il loro grosso nucleo (n), strettamente unite fra loro in lamine cellulari a direzione centripeta.
Fonte: Istituto di anatomia umana normale università degli studi di Bologna
Articolo creato il 23 febbraio 2012.
Ultimo aggiornamento: vedi sotto il titolo.
