Granulocitopoiesi
Il processo di differenziazione dei granulociti è caratterizzato da:
- Perdita graduale della basofilia citoplasmatica dovuta alla diminuzione dei ribosomi.
- Comparsa e progressivo accumulo di granulazioni specifiche (neutrofile, acidofile o basofile) che consentono di distinguere le tre sottolinee differenziative dei granulociti; la formazione dei granuli specifici comprende le caratteristiche fasi dei processi secretori: sintesi dei costituenti chimici sui ribosomi del reticolo endoplasmatico granulare nel complesso di Golgi, accumulo dei granuli specifici nell’interno dei vacuoli rivestiti di membrana.
- Modificazioni profonde del nucleo che comprendono la perdita dei nucleoli, la graduale condensazione della cromatina in grosse zolle eterocromatiche, il progressivo cambiamento della forma da rotonda a lobata o segmentata.
- Attività mitotica continua ed intensa fino allo stadio di mielocito che poi si arresta completamente.
- Attività ameboide che, assente nelle forme immature, compare solo nel granulocito definitivo il quale diventa così capace di migrare verso i sinusoidi e di attraversare la parete.
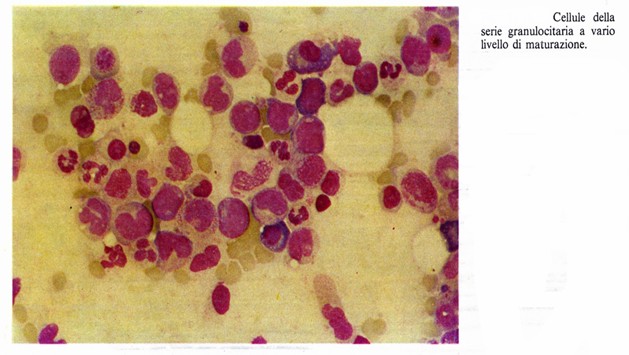
Le fasi di maturazione della granulocitopoiesi sono le seguenti:
Il mieloblasto rappresenta il primo stadio di maturazione riconoscibile morfologicamente. È una cellula relativamente piccola (12-20 µm di diametro) con un grosso nucleo eucromatico contenente 2-4 nucleoli ed un citoplasma basofilo ricco di ribosomi e privo di granulazioni.
Il promielocito rappresenta lo stadio di maturazione caratterizzato dalla comparsa delle granulazioni che consente di distinguere i pro mielociti neutrofili, basofili ed acidofili. Contemporaneamente al progressivo accumularsi di granuli, la basofilia citoplasmatica si attenua fino a scomparire. Nei promielociti neutrofili lo ialoplasma diventa leggermente acidofilo. Il nucleo si appiattisce e comincia a presentare una lieve intaccatura che delimita una zona citoplasmatica chiara occupata dal complesso di Golgi e dai centrioli. La cromatina si condensa e scompaiono i nucleoli. Il promielocito è più grande del mieloblasto (diametro di 15-20 µm) per il progressivo accumulo dei granuli. La cellula si divide attivamente.
Nei pro mielociti neutrofili la comparsa dei granuli primari (o azzurrofili) precede quella dei granuli secondari o specifici. I primi, più densi e caratterizzati dalla positività alla reazione istochimica per la perossidasi, fanno la loro comparsa allo stadio di promielocito ed originano dalla faccia concava del complesso di Golgi. I granuli specifici sono prodotti più tardivamente, durante lo stadio successivo del mielocito, e si originano dalla faccia convessa del complesso di Golgi. Nello stadio di promielocito l’attività perossidasica è rilevabile non soltanto nei granuli primari ma è diffusa a tutto l’apparato secretorio, cisterne del reticolo endoplasmatico e cisterne del complesso di Golgi. Quando nello stadio successivo la produzione dei granuli si arresta, la reazione perossidasica scompare dagli organuli secretori e permane nei granuli.
Nei granulociti acidofili e basofili i primi granuli specifici, ancora immaturi, compaiono in questo stadio.
Il mielocito ha un volume cellulare che si riduce e una basofilia citoplasmatica che scompare completamente; nei mielociti neutrofili ed acidofili, anzi, è sostituita da una debole acidofilia. Le granulazioni riempiono tutto il citoplasma ed aumenta la condensazione nucleare. Il nucleo si appiattisce ulteriormente e si accentua la sua intaccatura. Il complesso di Golgi è ben sviluppato, circonda i centrioli ed occupa una posizione paranucleare in vicinanza dell’intaccatura del nucleo. Sono presenti scarsi elementi di reticolo endoplasmatico granulare e numerosi mitocondri già riscontrabili negli stadi precedenti. La cellula presenta ancora attività mitotica.
Nei mielociti neutrofili sono presenti due tipi di granuli: primari (o azzurrofili), più densi e positivi alla reazione per la per ossidasi e contenenti varie idrolasi acide (lisosomi), e secondari (o specifici), più piccoli, meno densi agli elettroni e negativi alla reazione per la per ossidasi, che fanno la loro prima comparsa in tale stadio. Via via che si accumulano nel citoplasma, i granuli specifici diventano l’organulo prevalente (80% delle granulazioni citoplasmatiche). I granuli eosinofili presentano lievi variazioni durante la maturazione, sono di diametro maggiore dei granuli primari e un numero sempre maggiore di essi sviluppa nel suo interno un cristalloide. I granuli basofili non presentano modificazioni rilevanti durante la granulopoiesi.
Il metamielocito è caratterizzato dall’accentuazione dell’intaccatura nucleare di modo che il nucleo si presenta incurvato o reniforme. La divisione cellulare e la sintesi dei granuli citoplasmatici si arrestano e, come conseguenza, il complesso di Golgi e il reticolo endoplasmatico granulare si riducono di dimensioni e diventano inattivi.
Nei metamielociti neutrofili le due classi di granuli (granulazioni azzurrofile positive alla perossidasi, più grandi e più dense agli elettroni e granuli specifici negativi alla perossidasi, più piccoli e meno densi) sono tra loro nel rapporto 1:4.
Il granulocito maturo presenta un nucleo più deformato che diventa lobato o segmentato. Come il metamielocito è inattivo mitoticamente e non elabora i granuli citoplasmatici. Esso è una forma terminale incapace di ulteriore differenziazione.
L’attività ameboide compare solo al termine del processo maturativo rendendo così la cellula capace di attraversare la parete dei sinusoidi. Contemporaneamente nel citoplasma si osservano i movimenti caratteristici dei granuli (ciclosi) e si sviluppano filamenti actino-simili dello spessore di 5 nm. L’assenza di attività motoria della cellula negli stadi precedenti è apparentemente il meccanismo che previene la liberazione in circolo delle cellule immature.
Il processo di maturazione completo fino alla liberazione del granulocito maturo nella circolazione sanguigna impiega circa 14 giorni, di cui 7,5 per la fase mitotica dal mieloblasto al mielocito e 6,5 per la fase post-mitotica dal metamielocito al granulocito maturo. Gli elementi maturi rimangono nel circolo sanguigno per una piccola frazione del loro ciclo totale di vita (8-12 ore, con una vita media di 6 ore) e passano rapidamente nel tessuto connettivo dove sopravvivono per altre 24-48 ore prima di degenerare ed essere fagocitati dagli elementi del sistema reticolo-istiocitario.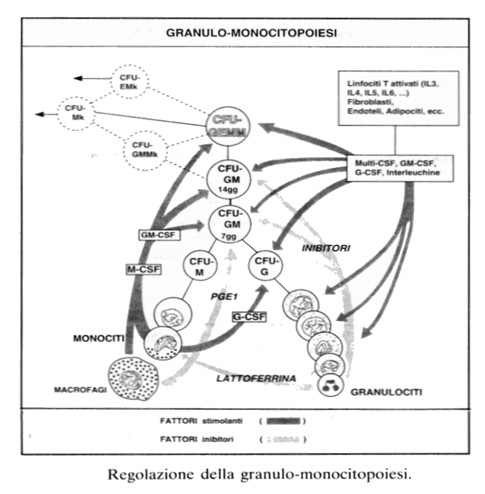
I granulociti, a differenza degli eritrociti ed analogamente ai monociti, devono essere quindi considerati più propriamente come cellule del connettivo che si servono del sangue come veicolo di transito dal luogo di produzione alla sede definitiva.
Sembrerebbe, infine, che nel midollo osseo si costituisca una riserva di granulociti maturi che possa essere rapidamente mobilizzata in casi di richieste funzionali.
Come la monocitopoiesi, la granulocitopoiesi è regolata da agenti stimolanti chiamati genericamente Fattori Stimolanti le Colonie (CSF), di origine diversa, che agiscono sia sui progenitori più immaturi (CFU-GEMM, CFU-GM) sia, insieme a fattori più specifici (G-CSF e M-CSF) sui progenitori commissionati per la sola granulo poiesi (CFU-G) e per la sola monocito poiesi (CFU-M). Questi agenti non sono altro che citochine, di natura glicoproteica, prodotte da monociti, macrofagi, linfociti attivati, ecc.. Il GM-CSF è anche chiamato IL3 prodotta da linfociti attivati. Essa agisce anche su altri progenitori emopoietici (CFU-Mk, BFU-E, ecc.).
La regolazione della granulo-monocitopoiesi avviene anche per la presenza di fattori a funzione inibitoria prodotti, ad esempio, dai granulociti (caloni, lattoferrina), oppure da macrofagi, come prostaglandine (PG1). Questi inibitori agiscono sui progenitori e precursori a livelli maturativi diversi e contribuiscono al mantenimento fisiologico della popolazione granulo-monocitaria.
Articolo creato il 6 marzo 2010.
Ultimo aggiornamento: vedi sotto il titolo.
