Recettore glicinergico (o della glicina) A
Così chiamato per distinguerlo dal recettore della glicina B che è un sito allosterico del recettore del glutammato di tipo NMDA, il recettore glicinergico (o della glicina) A (GlyRA) appartiene alla famiglia dei recettori-canale anionici (permette il passaggio di Cl– dall’esterno all’interno della membrana secondo gradiente 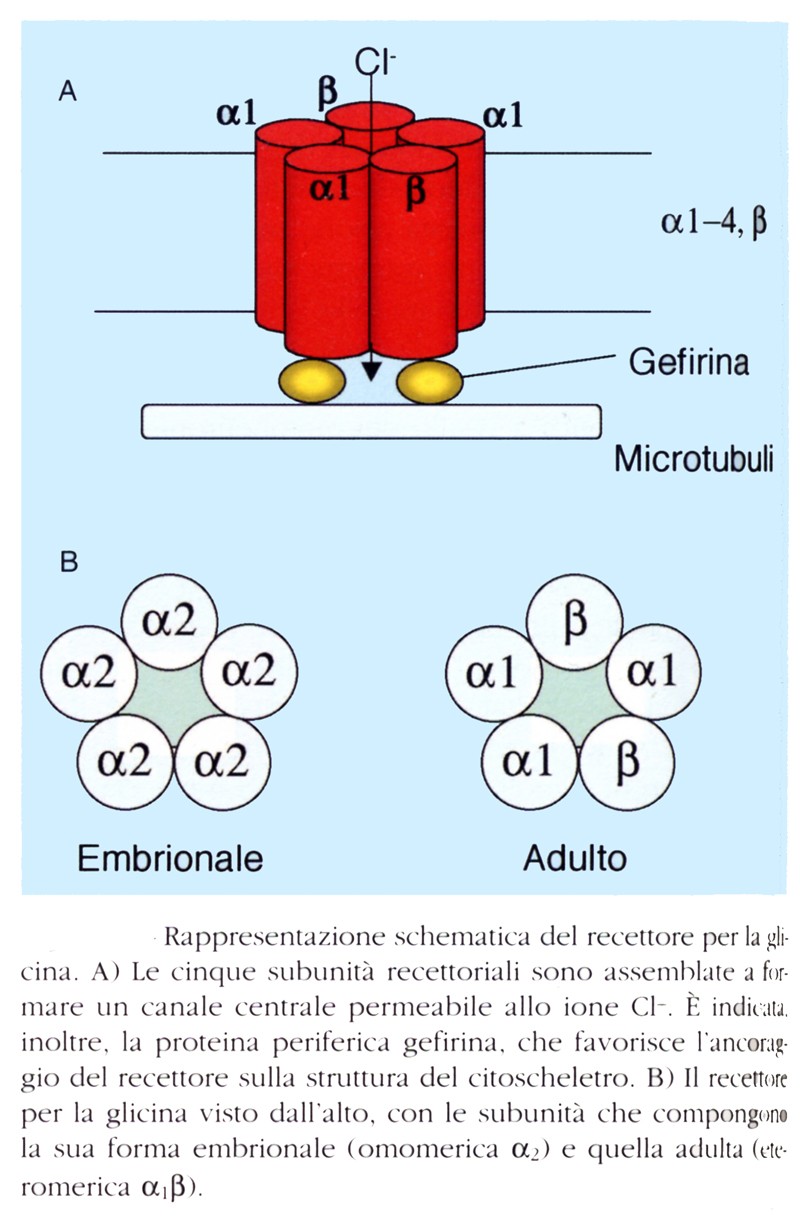 elettrochimico) nonché alla classe I (insieme al recettore colinergico nicotinico, al recettore del GABA di tipo A e al recettore della serotonina di tipo 3), quindi è un pentamero.
elettrochimico) nonché alla classe I (insieme al recettore colinergico nicotinico, al recettore del GABA di tipo A e al recettore della serotonina di tipo 3), quindi è un pentamero.
Tale tipo di recettore, sito all’interno di interneuroni brevi nella sinapsi laterale simmetrica a livello del midollo spinale, controlla la scarica dei motoneuroni delle corna anteriori del midollo spinale. Una caratteristica di questi recettori è che la composizione in subunità può cambiare sia durante lo sviluppo che nell’adulto. Il recettore di tipo A è costituito da subunità del tipo α1.
Il recettore della glicina è un complesso macromolecolare composto da una combinazione di due subunità omologhe α (α1-3) e β. La struttura quaternaria di questo recettore è costituita da cinque subunità disposte in maniera pseudosimmetrica a formare un canale ionico centrale permeabile agli ioni cloro. Le subunità α e β, come nel caso di quelle del recettore GABAA e del recettore colinergico nicotinico, presentano 4 domini transmembrana (TM1-TM4), un dominio amino-terminale extracellulare glicosilato, un ampio segmento citoplasmatico che collega TM3 e TM4, che contiene siti che possono essere fosforilati, e un dominio carbossi-terminale citoplasmatico. In aggiunta, è presente la gefirina, una proteina periferica altamente idrofila, localizzata a livello citoplasmatico in prossimità delle membrane postsinaptiche, la cui funzione è di cruciale importanza nell’addensamento (o clustering) dei recettori per la glicina a livello della membrana sinaptica, fungendo da elemento di ancoraggio con gli elementi del citoscheletro.
Quando il recettore è costituito da subunità α1, il recettore è attivato da glicina, taurina e β-alanina, mentre è bloccato da stricnina e picrotossina. Quando è costituito da subunità α2 e α3, invece, il recettore non è attivato da taurina e β-alanina.
Questi recettori sono altamente espressi nel midollo spinale, nel tronco encefalico, nell’ippocampo e nell’ipotalamo, mentre sono poco espressi nelle aree cerebrali superiori. Ancora, le subunità del recettore sono espresse anche nella retina, nelle ghiandole surrenali, nel rene, nel fegato e nelle cellule spermatiche.
La funzione di questi recettori è permettere l’apertura di canali al cloro sui neuroni postsinaptici in modo che questo ione entri nella cellula, iperpolarizzi la membrana e riduca così la possibilità che si sviluppi un potenziale d’azione.
Tuttavia, durante le fasi iniziali dello sviluppo, la glicina può agire come neurotrasmettitore eccitatorio; ciò è dovuto ad un valore più positivo del potenziale di equilibrio del cloro nei neuroni embrionali. In questi neuroni, quindi, la glicina attiva un’uscita di ioni cloro dalla cellula che induce una depolarizzazione della membrana e la conseguente apertuta di canali del calcio voltaggio-dipendenti di tipo L disposti sul corpo cellulare. Dopo la nascita, tuttavia, il potenziale di equilibrio del cloro si riassesta su valori più negativi e iperpolarizzanti, in conseguenza dell’espressione sulla membrana neuronale dello scambiatore K+-Cl–.
Articolo creato il 5 marzo 2010.
Ultimo aggiornamento: vedi sotto il titolo.

Potrebbe anche piacerti:
