Recettori dei fattori di crescita
I recettori dei fattori di crescita, come il recettore dell’insulina e i recettori delle citochine, fanno parte dei recettori ad attività tirosin-chinasica intrinseca (TRK).
Essi condividono una struttura di base costituita, nella maggior parte dei casi, da una singola catena polipeptidica che attraversa una sola volta la membrana plasmatica. Dal punto di vista funzionale, tali recettori sono organizzati in diversi domini funzionali:
- Il dominio extracellulare comprende il sito di legame per il fattore di crescita, presenta diverse combinazioni di motivi strutturali e rappresenta la parte più variabile del recettore. Sulla base di queste differenze i recettori per i fattori di crescita possono essere raggruppati in diverse sottofamiglie.
- Il dominio transmembrana è costituito da circa 25 aminoacidi idrofobici che attraversano il doppio strato lipidico e funge da connessione tra il dominio extracellulare e quello intracellulare.
- Il dominio iuxtamembrana comprende una cinquantina di aminoacidi, separa il dominio transmembrana da quello catalitico ed è sede di importanti funzioni regolatorie. È una regione ben conservata tra recettori appartenenti alla stessa sottofamiglia ed è coinvolta nella modulazione della trasduzione del segnale da parte di segnali extrarecettoriali.
- Il dominio catalitico, responsabile dell’attività tirosin-chinasica, è formato da 250 aminoacidi e ha un’elevata omologia fra tutti i recettori per i fattori di crescita.
- La coda C-terminale ha lunghezza e funzioni variabili e rappresenta il segmento che, nel recettore attivato, lega i trasduttori intracellulari del segnale.
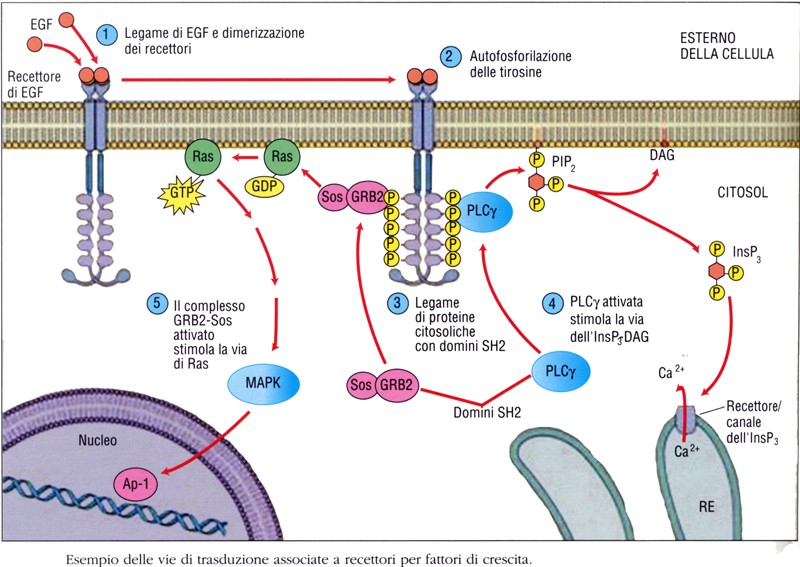
La maggior parte dei recettori per i fattori di crescita sono omodimeri e possiedono un meccanismo classico secondo il quale il fattore si lega al dominio extracellulare, evento a cui fa seguito la dimerizzazione del recettore. Il legame con il recettore induce un cambiamento conformazionale del dominio extracellulare che stabilizza le interazioni recettore-recettore.
La dimerizzazione stabilizza lo stato attivo della chinasi e porta in sua prossimità il dominio intracellulare di un altro recettore. Conseguentemente si verifica una transfosforilazione dei recettori a livello dei residui di tirosina situati sia all’interno che all’esterno del dominio catalitico. La transautofosforilazione del recettore può verificarsi anche esternamente al dominio catalitico, nel dominio iuxtamembrana e nella coda C-terminale.
La fosforilazione di questi residui porta alla formazione di siti di ancoraggio che vengono riconosciuti con alta affinità dai trasduttori citoplasmatici del segnale contenenti specifici domini.
I trasduttori intracitoplasmatici del segnale si legano con alta affinità ai recettori fosforilati per mezzo di specifici “moduli” contenuti all’interno della loro sequenza. Il più comune di questi è il cosiddetto dominio SH2 (da SRC homology region 2) che riconosce e lega corte sequenze aminoacidiche contenenti una tirosina fosforilata; un altro modulo è il modulo PTB (da phospho-tyrosine binding). L’associazione selettiva di trasduttori contenenti domini SH2 a specificità diversa determina quale via di trasduzione del segnale verrà attivata.
Esistono due classi di trasduttori provvisti di domini SH2:
- La prima classe è rappresentata da proteine che possiedono attività enzimatica propria, come la fosfolipasi C-g (PLC-g), la tirosin-chinasi codificata dal proto oncogene c-SRC (pp60src), le fosfotirosin-fosfatasi SHP1 e SHP2, l’attivatore della GTPasi (GAP) e i trasduttori del segnale attivatori trascrizionali (STAT).
- La seconda classe raggruppa trasduttori privi di attività catalitica intrinseca. Questi possiedono un ruolo di subunità regolatrici di molecole con attività enzimatica citoplasmatiche; esempi sono: Grb2, Shc e la subunità non catalitica della fosfatidil-inositolo-3-chinasi (PI3K).
Articolo creato il 6 marzo 2010.
Ultimo aggiornamento: vedi sotto il titolo.


Potrebbe anche piacerti:
